
tRNA-wobble位ウリジン修飾の生物学的重要性に関する研究
大阪医科薬科大学 生化学教室 准教授
yumi.nakai(at)ompu.ac.jp
[ 真核生物tRNA-wobbleU34修飾とは ]
tRNA塩基の転写後修飾はこれまでに200種類以上の修飾基とそれぞれに対する修飾関連タンパク質群が同定されています。中でも 翻訳に直接関わるアンチコドンとその周辺(アンチコドンループ)にはそれぞれのtRNAに特有の多様な塩基修飾がありtRNAが適切な翻訳機能を果たすためにこれらの塩基修飾が重要であることを示しています。これらは、tRNAの安定性の維持の他、翻訳精度の向上や翻訳速度の適正化に重要であることが最近の研究でわかってきました。
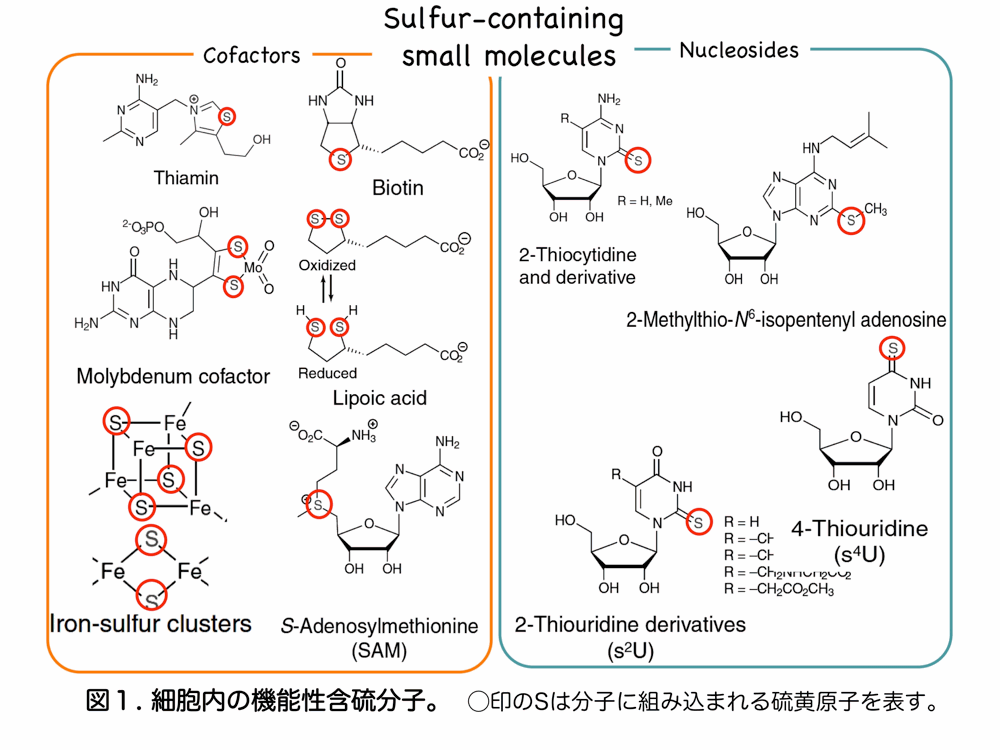
多細胞真核生物では、細胞質のtRNALys(UUU)、 tRNAGlu(UUC)、およびtRNAGln(UUG)の最初のアンチコドンウリジン塩基(wobbleU34)は、転写後修飾によりメトキシカルボニルメチル基の添加と、第2炭素に結合している酸素から硫黄への置換を受けて5-メトキシカルボニルメチル-2-チオウリジン(mcm5s2U)を形成しており(図1) (ここではtRNA-wobbleU34修飾とよぶことにします)、この修飾は適切なコドン-アンチコドンペアの形成に寄与すると考えられています。mcm5s2U形成に関わるタンパク質群はmcm5形成には800kDaを超える巨大なタンパク質複合体Elongatorとメチル基転移酵素を必要とします。一方、s2形成は、ウリジンの2位炭素に結合している酸素を硫黄にわざわざ置換するという複雑な反応であり、細胞内のシステインから硫黄を引き抜くシッフ塩基含有酵素Nfs1や、硫黄運搬酵素Cnx5、チオカルボキシル基を形成するユビキチン様硫黄運搬酵素Urm (ヒト・酵母ではURM1, シロイヌナズナではURM11とURM12) などが、tRNAチオ修飾付加酵素とともに働くといった一連の反応を経て付加されます。この様な複雑な多段階反応で付加される2箇所の異なる塩基修飾を併せ持つtRNA-wobbleU34修飾は多くの真核生物にみられ、またその修飾基形成タンパク質群もよく保存されていて、生物にとってこの修飾が重要であることを示しています。
[tRNA-wobbleU34修飾欠失が生物にもたらすものは?]
動物の細胞においてtRNA-wobbleU34修飾形成に関わる修飾酵素の異常や欠失は神経変性疾患や免疫の低下、あるいは癌細胞のタンパク質翻訳効率変化と関連することが報告されています。一方、tRNA-wobbleU34修飾を構成する2つの修飾形成経路を同時に欠損させると、マウスなどの哺乳動物やショウジョウバエでは致死性を引き起こすことが知られています。また、部分欠失でも細胞増殖や細胞分化の正常過程に障害が出ます。構造的に見るとtRNA-wobbleU34修飾は特にYAAコドン(YはA,C,Gのいずれかの塩基)翻訳時の3番目(wobble位)のコドンーアンチコドン対合(A-U)の安定化に重要であり、このコドンの翻訳を適切に行うことは細胞機能に関わる多くのタンパク質発現に全般的に重要であることは容易に推察できます。しかし、その修飾欠失で致死に至る理由として、細胞増殖や分化など生命機能を支えるためのプログラムのどのような作用点に特に強く機能しているのかは明らかになっていません(図2)。
tRNA-wobbleU34修飾が実際に多細胞真核生物の生命維持に関わるどのような機能に重要なのかを知るためには、tRNA-wobbleU34修飾欠失でも致死ではないが、何らかの重篤な影響が出るような多細胞真核生物を調べることが1つの手がかりとなります。そこで目をつけたのが植物です。植物は酵母などの単細胞真核生物と異なり、動物同様、細胞が増殖と分化のステップをへて組織や器官といった機能分化した複雑な細胞集団を形成し、それらの機能のオーケストレーションが個体を作り上げています。この過程で動物にはなく植物にあるのが、植物細胞(特に幹細胞)の遺伝子発現の柔軟性です。植物は、一定の決まった大きさで成長が止まる動物の「有限成長」とは対照的に、茎頂分裂組織や根端分裂組織にある幹細胞(メリステム)を永続的に維持してそこから器官・組織形成につながる細胞の増殖と分化を継続させる能力が動物と比べて極めて高いため、動物では致死に至る機能変異でもなんとか持ち堪えて個体を維持できる可能性があります。これは植物が動物と異なり動くことができない環境に適応するため獲得した性質であると言われますが、この点に着眼し、植物のtRNA-wobbleU34修飾欠損は、動物同様、生命機能に関わる遺伝子発現に深く影響するものの、致死にはならない可能性があると考えました。そこで、モデル植物のシロイヌナズナにおけるtRNA-wobbleU34修飾欠失株を作成し調べたところ、期待どおり、Elongator活性サブユニットELO3/ELP3と遺伝子重複で2つ生じている硫黄運搬タンパク質URM11, URM12の3つの遺伝子変異株はtRNA-wobbleU34修飾欠失でありながら致死にならないことを見出しました。さらに興味深いことに、tRNA-wobbleU34修飾欠失は植物において致死とはなりませんが、顕著な生育遅延と様々な形態異常を呈することを見出しました。このことは、植物のさまざまな発生・細胞分化・成長過程において、tRNA-wobbleU34修飾の欠損の影響が特に大きく現れる作用点が幾つか存在する事を示しています。それゆえ、動物ではその欠損が致死になってしまうため調べるのが難しいtRNA-wobbleU34修飾について、植物を用いる事で、欠損の影響が特に大きく現れる局面に着目し、その原因の詳細を解明することが可能になる、と考えています。加えて、植物tRNA-wobbleU34修飾欠損が引き起こす細胞機能障害の作用点に対応するものが動物ではどのようなものなのかを明らかにすることにより、tRNA-wobbleU34修飾がヒトを含む多細胞真核動物の生命機能維持にいかに関わるのかを解明することを目指しています。